Under utdelingen av nobelprisen i medisin og fysiologi i Stockholm i 1962, manglet én person på podiet; den unge kjemikeren Rosalind Franklin, som hadde fremstilt bilder av spiralstrukturen lenge før de tre som mottok hederen. Hadde hun levd, ville hennes innsats blitt trukket frem og verdsatt langt tidligere enn den ble.

Nobelprisen i medisin for 1962 ble delt av James Watson, Francis Crick og Maurice Wilkins (hhv. nr. 5, 3 og 1 fra venstre) for oppdagelsen av DNA-molekylets struktur. De øvrige prisvinnerne på bildet er Max Perutz i kjemi (nr. 2 fra venstre), John Steinbeck i litteratur (nr. 4) og John Kendrew i kjemi (nr. 6). Foto Bettmann, Corbis/Scanpix

Den avdøde kjemikeren Rosalind Franklin, som med sine eiendommelige krystallografibilder av DNA hadde banet vei for dechiffreringen av molekylets struktur, fikk ikke del i nobelæren da prisen ikke deles ut posthumt. Verken Watson eller Crick nevnte henne i sine taler. I det 29-siders manuset til Wilkins tale, er det bare én henvisning til hennes arbeid. Foto Novartis Foundation.
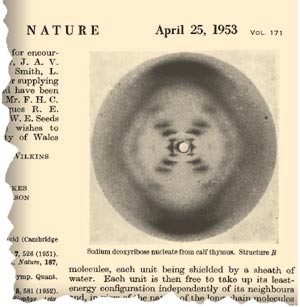
Faksimile fra Franklin & Goslings artikkel i Nature med det berømte bildet av den såkalte B-formen av DNA.
På podiet stod James Watson, Francis Crick og Maurice Wilkins side om side og mottok nobelprisen i medisin og fysiologi. Ni år før ble DNA-æraen innledet med Natures publisering av Watson & Cricks artikkel om DNA-molekylets spiralstruktur 25. april 1953 (1).
Kort artikkel, rikt innhold
På litt over én side og med henvisning til seks andre publikasjoner, argumenterer de to forskerne for hvorfor DNA, deoksyribonukleinsyre, nødvendigvis må ha form som en dobbel heliks bestående av to parallelle, sammenvevde kjeder av kjernesyrer. Modellen ble understøttet av krystallografiske data publisert i samme nummer av Nature, av henholdsvis Wilkins og medarbeidere (2) og Franklin & Gosling (3).
Fra første stund ble det klart at spiralstrukturen også røpet en mekanisme for hvordan arvestoffet lar seg kopiere, eller replikere, til et nærmest uendelig antall nye, identiske molekyler. Dette var også tema i en Watson & Cricks oppfølgerartikkel samme år (4). Nå var det ikke lenger noen tvil om at det var DNA, og ikke proteinene, som var arvens materielle substrat og bæreren av cellens genetiske informasjon; det som sjargongmessig ble kalt for «the stuff of life».
Livets hemmelighet
28. februar 1953 var dagen da amerikanske James Watson (1928–) og engelske Francis Crick (1916–), som begge arbeidet ved Cavendish-laboratoriet ved University of Cambridge, for første gang kunngjorde at de hadde lyktes i å konstruere en ikonisk modell av DNA-molekylets tredimensjonale struktur.
– Vi har avdekket livets hemmelighet, forkynte Watson da han samme ettermiddag besøkte Cambridge-puben The Eagle. Men det var ingen der som skjønte hva den frittalende amerikaneren snakket om. I 1953 var det heller ikke noe offentlig blest rundt DNA-oppdagelsen, og det skulle gå flere tiår før genteknologien fullt ut kunne utnytte potensialet som lå i arvestoffstruk-turen. Like fullt var det et gjennombrudd av dimensjoner, og Time-skribenten Robert Wright uttrykte det som «resultatet av en unik resonans mellom to ulike forskerpersonligheter, som gjennom felles innsats og en forening av ideer, mot og ambisjoner utfordret sin tids akademiske oppfatninger og dogmer» (5).
Crick var utdannet fysiker fra London og Cambridge, men hadde penset over på kjemi og biologi på grunn av sin fascinasjon for «linjen mellom det levende og det ikke-levende». Han hadde ennå ikke disputert, i motsetning til den 12 år yngre Watson, som var biolog med doktorgrad på bakteriofager fra universitetene i Chicago og Indiana i USA.
Watson la vekk sin opprinnelige interesse for fugler til fordel for virus, og drog til Europa i 1950 for å tilegne seg mer biokjemisk kunnskap om arvestoffstudier. Han kom etter hvert i kontakt med det ledende engelske forskningsmiljøet i Cambridge, der han møtte Francis Crick.
Sjelden allianse
At DNA var det genuine arvestoffet, var allerede i 1943 bevist gjennom bakteriologiske studier gjennomført av Oswald Avery, Colin MacLeod og Maclyn McCarty (6). De tre forskerne ved Rockefeller-instituttet i New York hadde vist at virulente egenskaper hos pneumokokker kunne overføres fra døde, sinte stammer til levende, ufarlige bakterier. Det overførte materialet var nettopp deoksyribonukleinsyre.
I Norge ble dette funnet kommentert av den unge bakteriologen Sverre Dick Henriksen i en artikkel i Tidsskriftet i 1947 (7). «Det som er nytt, og som kanskje kan få stor betydning for andre felter av biologien, er at det ser ut som om det er mulig å innføre nytt arvestoff i bakteriekjernen, og derved fremkalle mutasjoner i forut bestemt retning,» skrev Henriksen, som også antydet at Averys oppdagelse av DNA som arvematerialet var verdig en nobelpris i medisin.
Da James Watson på en konferanse i Napoli noen år senere, iakttok en vag og nesten spøkelsesaktig avbildning av DNA, fremstilt ved røntgenkrystallografi, ble han nærmest besatt av tanken på å løse gåten rundt molekylets struktur. «Det var umulig å bli kvitt ideen om en potensiell nøkkel til livets hemmelighet,» skrev han senere i memoarboken The double helix, som kom ut i 1968 (8). «Muligheten til å bli berømt, fristet meg mye mer enn utsiktene til å ende opp som en tilstivnet akademiker som aldri hadde våget et tankesprang.»
Watson hadde ikke bare ord på seg for å være flink, han var også kjent som en ubeskjeden og høyrøstet fyr. Amerikanerens erklærte målsetting om å vinne DNA-kappløpet i konkurranse med forskere som Linus Pauling og Erwin Chargaff, slik det fremgår av boken hans, ble imidlertid ikke applaudert av makker Francis Crick, som i et tidsskriftinnlegg i 1974 gikk i rette med den økende strømmen av beskyldninger fra flere hold om at ære og prestisje hadde vært den sentrale beveggrunnen for de to forskerne (9). «Min erindring var at vi, for å si det helt enkelt, var heltent på å løse DNA-gåten,» uttalte han senere (5).
Uansett hvilke motiver de hver for seg måtte ha hatt, delte Watson og Crick den samme forskertrangen. Når de så å si kveilet sammen sin fascinasjon for DNA-spiralen i laboratoriene ved Cambridge, skapte det en allianse av de sjeldne i forskningsverdenen.
Franklins DNA-bilde
Likeså ubestridelig som at Watson og Crick var arkitektene bak den doble DNA-heliksen, er det at andre vitenskapsfolk hadde gjorde mye av forarbeidet til nøkkeloppdagelsen. En stråle av glansen falt på den norske kjemikeren Sven Furberg (1920–83), opphavsmannen til en av de seks referansene i Watson & Cricks litteraturhenvisninger (10). Furbergs røntgenstudier av kjernesyrer hadde ført ham til Oxford, der han arbeidet videre med krystallografiteknikken i det samme miljøet som Watson og Crick hentet sine observasjoner fra (11).
Oppdagerne fikk sin belønning i 1962 da de ble hedret med nobelprisen i medisin og fysiologi, en pris de delte med biofysikeren Maurice Wilkins (1916–) som arbeidet med DNA ved King’s College i London. Det var likevel et annet arbeid som for alvor hadde overbevist Watson om at DNA-molekylet bestod av en dobbel heliks. Ved samme institutt som Wilkins, hadde kjemikeren Rosalind Franklin (1920–58) laget enestående røntgendiffraksjonsbilder av DNA. Franklin, som var førsteforfatter på den ene av de to nevnte parallellpublikasjonene i Nature (3), var 33 år gammel i oppdagel-sesåret og arbeidet som biofysiker ved King’s College.
Ved hjelp av krystallografiteknikken som gjorde det mulig å kartlegge atomenes plassering i krystaller, hadde hun allerede konstatert at DNA-molekylets fosfatgrupper lå på utsiden av dobbelspiralen og at DNA eksisterte i to former (12). Medio februar, bare et par uker før Watson og Crick røpet sin oppdagelse, skrev hun i notatboken sin at molekylet bestod av to kjeder med nukleinsyrer (13, 14).
I ettertid kom det for dagen at Maurice Wilkins mislikte Franklin og anså henne for å være en gryende feminist (5). Ifølge Watson var det kjølige forholdet gjensidig, og en direkte årsak til at Wilkins helt uoppfordret på nyåret 1953 viste ham et av Franklins beste DNA-bilder som ennå ikke var publisert. Dette gjorde han helt uten hennes samtykke og uten at hun visste noe som helst. «Da jeg fikk se bildet, kunne jeg ikke annet enn måpe,» innrømmet Watson senere (8). Han hadde sett lyset; Franklins dårlig bevarte hemmelighet var for ham blitt en åpenbaring av spiralstrukturen.
Ulike versjoner
Rosalind Franklins karriere ble kortvarig. Hun døde av ovarialcancer i 1958, bare 37 år gammel, og rakk verken å elaborere sin DNA-teori eller forsvare sin innsats i arbeidet med spiralmodellen. Den offisielle forklaringen på at hennes navn ble utelatt ved nobelpristildelingen i 1962, har vært at prisen ikke kan tildeles etter døden. Det faktum at Watson og Crick unnlot å referere til hennes arbeid i artikler og nobelforedrag, er blitt sett på som en gedigen forbigåelse. I ettertid vedgikk de begge å ha sett Franklins krystallografibilder, men Watson nedtonet likevel implikasjonene av dette. Franklin-biografen Brenda Maddox kaller hele historien for et «kroneksempel på den rådende mysogyni i forskningens verden» (14).
Således foreligger det to diametralt forskjellige versjoner av DNA-kappløpet som fant sted forut for våren 1953; den ene fortalt av James Watson i hans memoarbok, og den andre formidlet av Sayre (15) og Maddox (16) i biografiene om Rosalind Franklin fra henholdsvis 1975 og 2001. Mens Watson tegner et svært lite flatterende bilde av Franklin, både som forsker og person, blir hun av de to forfatterne portrettert som en begavet, kultivert og sky person som følte seg ensom og motarbeidet ved mannsbastionen King’s College.
«Dersom Franklin hadde levd, ville det vært umulig å forfordele henne ved nobeltildelingen, fordi det var hun som stod bak flere av nøkkelforsøkene,» har Francis Crick sagt offentlig. Men hennes bidrag slutter med det (5); det er også kommet for en dag at hennes kritikk av Watson og Cricks tidlige DNA-modeller hadde sendt parhestene tilbake til tegnebrettet. Og av egenhendige notater har det fremkommet at hun selv hadde som mål å løse gåten rundt DNA-strukturen. Hun hadde skjønt at molekylet måtte ha en eller annen spiralform, men hun unnlot å benytte knepet som Watson og Crick så til overmål godt lyktes med: å fikle med store, tredimensjonale modeller. Det var det som førte dem til suksess.
Genetisk kode
Det var Watson som fikk DNA-puslespillet på plass, da han ved bruk av pappmodeller pønsket ut hvorledes de fire nitrogenbasene adenin (A), tymin (T), cytosin (C) og guanin (G) danner par som føyer de to enkelttrådene i DNA-molekylet sammen til en form som likner på en vridd stige. Watson så at A og T, forbundet med to hydrogenbindinger, har nøyaktig samme konfigurasjon som baseparet C og G. Det gikk opp for ham at disse unike baseparene utgjør trinnene i den vridde DNA-stigen.
Crick, på sin side, klekket ut det sentrale konseptet om samhørighet eller komplementaritet mellom A og T og mellom C og G. Det gjorde han på basis av funnene til Erwin Chargaff, som viste at DNA inneholder nøyaktig like mye A som T og C som G. Det eiendommelige ved konseptet om komplementaritet var at det var nøkkelen til å forstå hva som skjer på molekylplanet når en kroppscelle deler seg og kromosomene dupliseres: Kopieringen eller replikasjonen av DNA innebærer at de to enkeltkjedene skiller lag og hver for seg danner en støpeform (templat) for en ny kjede. Enhver sekvens av det genetiske alfabetet i DNA, for eksempel CAT, vil dermed inngå i parvise forbindelser med en komplementær baserekkefølge, i dette tilfellet GTA. Ettersom basene danner spesifikke par, blir de to nye dobbeltspiralene identiske med originalen, og dattercellene får hvert sitt sett av like kromosomer.
I 1960-årene ble det klart at baserekkefølgen i DNA kontrollerer cellenes proteinsyntese via et mellomledd, som er budbringermolekylet ribonukleinsyre (RNA). Biokjemikere kartla nøyaktig hvordan de kjemiske bokstavinstruksene i DNA lovmessig ble oversatt til spesifikke aminosyrer som utgjorde byggesteinene i proteinene. Prinsippene for denne oversettelsesprosessen, kalt translasjon, ble spikret fast av Crick, Brenner og Barnett i 1961 (17), og fikk betegnelsen den genetiske kode. Fire år tidligere hadde Francis Crick formulert sammenhengen mellom kjernesyrer og proteiner i sitt velkjente sentraldogme (18), som slo fast at «DNA lager RNA som lager proteiner». Med andre ord, at det genetiske budskapet går bare én vei og at DNA følgelig er arvestoffet.
I skyggen av suksess
Den eiendommelige synergien mellom Watson og Crick blir ofte trukket frem når spørsmål reiser seg om hva som var nøkkelen til deres suksess, eller hvorfor ikke Rosalind Franklin kom først i mål i DNA-kappløpet. Det som er klart, er at hun manglet allianser og støttespillere i sitt eget forskningsmiljø, som var dominert av menn og av diskriminerende regler; blant annet hadde ikke kvinnelige ansatte adgang til enkelte av fakultetets oppholdsrom som var forbeholdt menn. Det var på disse arenaene at det uformelle fellesskapet og mye av den vitenskapelige diskusjonen utspant seg.
Utestengt fra de indre gemakker ved King’s College, ble Franklin stadig mer isolert. Hennes mistrivsel kulminerte, og våren 1953 var hun vel så opptatt av å komme seg vekk og over til et nytt sted, som å kjempe alene mot en mannsbastion av forskere. Hun takket ja til en post ved Birkbeck College i London, der hun også skiftet forskningstema i retning av virusstudier.
Overraskende nok, gav hun aldri uttrykk for faglig bitterhet mot Watson, Crick og Wilkins. I stedet støttet hun lojalt opp om arbeidet deres. Men så fikk hun heller aldri kjennskap til at de hadde sett resultatene hennes; det ble først avdekket ti år etter hennes død. Den dag i dag er det fortsatt få lærebøker som nevner hennes formidable innsats i beretningen om DNA-kappløpet, og derfor er det med rette blitt sagt at hun var vitenskapskvinnen som ble skrevet ut av DNA-historien (14).
James Watson og Francis Crick, på sin side, kastet mynt om hvem av dem som skulle stå først i deres felles byline: Crick & Watson, eller Watson & Crick? Det ble amerikaneren som trakk det lengste strået. James Watson flyttet tilbake til USA og bygde opp DNA-laboratoriet i Cold Spring Harbor i New York, som etter hvert fremstod som et fyrtårn i internasjonal molekylærbiologi. I 1990-årene ledet han arbeidet med Humant genom-prosjekt, historiens største biomedisinske forskningsprosjekt som i store trekk ble fullført i 2001. Francis Crick fortsatte med DNA-studier i England i et par tiår før han tok spranget over til nevrofag og hjerneforskning ved Salk-instituttet i California.
Sentral nordmann
Den norske kjemikeren Sven Furberg (1920–83) var opphavsmannen til referanse nummer 2 i den klassiske artikkelen om den doble heliksen. Watson & Crick henviste til Furbergs målinger av nukleotidstrukturen, som også var tema for hans doktorgradsarbeid i London i 1949–52. Nordmannen var først ute med å foreslå en heliksstruktur for nukleinsyrene, og benyttet røntgenkrystallografi til studier av biologisk viktige molekyler. Han forlot imidlertid England allerede i 1953, til fordel for en dosentstilling innen kjemi ved Universitetet i Bergen. I 1956 etablerte han, på oppdrag fra UNESCO, et røntgenkrystallografisk laboratorium i Montevideo. Året etter ble han dosent ved Universitetet i Oslo, og i 1966 kjemiprofessor samme sted.

Sven Furbergs nukleotidstudier var en del av grunnlaget for oppdagelsen av DNA-strukturen. Foto Kjemisk institutt, Universitetet i Oslo
Stille DNA-debut
Oppdagelsen av DNA-strukturen hadde i første omgang ingen praktisk betydning for biologien og genetikken (1). På 1950- og 60-tallet hadde forskere sin fulle hyre med å finne ut hva genene bestod av rent fysisk, og hvordan de oppførte seg i levende organismer. Watson & Cricks modell innebar et paradigmeskifte; genene var ikke lenger bare punkter på kromosomene, men definerte informasjonsbiter i DNA-molekylet som kontrollerte cellens biologiske prosesser. Spørsmålet var hvordan disse informasjonsbitene var satt sammen og hvilke mekanismer som sørget for at genene ble uttrykt i den levende organismen.
Det gikk et par stillferdige tiår før DNA-oppdagelsen for alvor satte fart i arbeidet med å kartlegge arvestoffet. Det første store skrittet ble tatt da den genetiske kode ble avdekket i 1960-årene. Oppdagelsen av restriksjonsenzymer i 1960- og 70-årene gjorde det mulig å kutte opp DNA i bestemte segmenter. Metoder for å transformere bakterier til å produsere små DNA-biter, gav forskerne muligheten til å klone og karakterisere gener og genprodukter. Dette banet vei for rekombinant DNA-teknologi som åpnet for massefremstilling av legemidler, i første rekke insulin, veksthormon, hematopoietiske vekstfaktorer og koagulasjonsfaktorer. I 1980-årene ble polymerasekjedereaksjonen (PCR) tatt i bruk for å fremstille DNA, et prinsipp som nordmannen Kjell Kleppe var den første til å beskrive i 1971. I 1990-årene ble effektive sekvenseringsteknikker lagt til grunn for genkartleggingsarbeidet i Humant genomprosjekt (HGP), som i grove trekk ble sluttført i 2000. De siste årene har DNA-mikromatriseteknologi gitt håp om en mer nyansert sykdomsdiagnostikk i fremtiden, basert på genanalyser.
Nielsen TH, Monsen A, Tennøe T. Livets tre og kodenes kode – fra genetikk til bioteknologi. Oslo: Gyldendal, 2000.
1869 – Fritz Miescher påviser en syrlig substans i kjernen på hvite blodceller som han benevner som nuklein. Så oppdager han at nuklein består av protein og et stoff som får navnet nukleinsyre og senere DNA.
1919 – Phoebus Aron Levene antyder at DNA har en tetranukleotid-struktur der de fire basene er ordnet i rekkefølge og i sett på fire.
1928 – Frederick Griffith oppdager at et ukjent stoff i varmedrepte bakterier kan overføre egenskaper til levende nabobakterier, et fenomen han kaller transformasjon.
1938 – Rudolf Signer, Torbjørn Caspersson og Einar Hammersten anslår molekylvekten for DNA til mellom 500 000 og 1 million atomenheter. Levenes’ tetranukleotid må være et polytetranukleotid.
1944 – Oswald Avery, Colin MacLeod og Maclyn McCarty identifiserer det kjemiske stoffet bak Griffiths transformasjonsprinsipp som DNA, og fremholder dette som arvematerialet.
1949 – Erwin Chargaff slår fast at fordelingen av baser i DNA varierer mellom ulike arter; men at det bestandig er like store mengder av de to purinbasene adenin og tymin, og likeledes et en-til-en-forhold i mengden av de to pyrimidinbasene cytosin og guanin. Roger og Colette Vendrely, sammen med André Boivin viser at kjønnsceller har halvparten så mye DNA i kjernen som det kroppsceller har, et funn som samsvarer med reduksjonen i antall kromosomer og styrker teorien om at DNA er arvematerialet.
1950 – Sven Furberg foreslår at kjernesyrene i DNA danner en spiralform, men han antar at molekylet bare har én nukleotidkjede.
1951 – Rosalind Franklin visualiserer DNA-molekylet ved hjelp av røntgenkrystallografi. Franklins arbeid frembringer to hovedformer: den parakrystallinske B-formen og den krystallinske A-formen. James Watson får kjennskap til denne metoden gjennom Franklins samarbeidspartner ved King’s College i London, Maurice Wilkins. I Cambridge møtes Watson og Francis Crick, og bestemmer seg for å løse gåten rundt DNA-strukturen. De lanserer en trippelheliksmodell som avvises av Franklin, på basis av krystallografidata. I USA lykkes Linus Pauling med å dechiffrere strukturen til proteinet keratin, og røper sine planer om å kartlegge DNA-strukturen.
1952 – Al Hershey og Martha Chase demonstrerer at bakteriofager (virus som angriper bakterier) overfører DNA, og ikke protein, når de infiserer bakterier for å produsere og spre nye bakteriofager. Rosalind Franklin og Raymond Gosling fremstiller sitt berømte røntgenbilde av B-formen til DNA. Det oppstår samarbeidsproblemer mellom Franklin og Wilkins. Linus Pauling prøver å komme til England for å gjøre DNA-studier sammen med Wilkins, men nektes utreise fra USA, på politisk grunnlag.
1953 – I januar fremsetter Pauling sin teori om at DNA har tre nukleotidkjeder, men venter med å publisere. Watson og Crick avfeier Paulings modell. Wilkins viser Watson Franklins beste DNA-bilde, «photograph 51», som antyder en spiralstruktur. 8. februar får Watson og Crick innsyn i en rapport om DNA fra King’s College, og blir overbevist om at molekylet har to kjeder. De løser gåten rundt basenes plassering, og 28. februar ferdigstiller de modellen av den doble heliksen, som blir presentert i en artikkel i Nature 25. april. I 1953 publiserer også Franklin & Gosling, Wilkins, Seeds, Stokes, Wilson samt Jacobsen artikler som belyser DNA-strukturen.
1954 – George Gamow postulerer at DNA inneholder den kjemiske koden for proteinsyntesen.
1955 – Seymour Benzer beskriver i detalj genstrukturen i bakteriofager.
1957 – Francis Crick legger frem sekvenshypotesen og sentraldogmen om at genetiske beskjeder bare går én vei (fra DNA via RNA til protein).
1958 – Matthew Meselson og Franklin Stahl oppdager en replikasjonsmekanisme for DNA.
1959 – Arthur Kornberg og medarbeidere isolerer enzymet DNA-polymerase.
1960 – budbringermolekylet mRNA blir identifisert.
1961 – Marshall Nirenberg og Johann Heinrich Matthaei avdekker den første av i alt 64 genetiske tre-bokstavs-koder for proteinsyntesen. Grunnlaget for å dechiffrere den genetiske kode er nå etablert.
1962 – Crick, Watson og Wilkins får nobelprisen i medisin.
Kilder: Nature og Time Magazine
