Som student, for ikke så mange år siden, var jeg med på en kirurgisk forelesning om medfødte hjertelidelser. Mot slutten av en interessant leksjon spurte jeg om personer som vokser opp takket være avansert hjertekirurgi, har økt risiko for selv å få barn med hjertefeil. Noe unnvikende svarte professoren at det er en markant økning av slike defekter blant barn av hjerteopererte. I spontan nysgjerrighet spurte jeg hvordan han forholdt seg til at hans kirurgiske behandling produserte flere barn med hjertefeil. Professoren tok da to lange skritt opp i auditoriet, og til min overraskelse ble nysgjerrigheten kontant avvist med at slike spørsmål og menneskesynet det insinuerte, ikke hørte hjemme på hans forelesninger. I ettertid har jeg forstått at jeg her uforvarende hadde anvendt et vitenskapelig prinsipp med en formidabel evne til å provosere, en logikk som har snudd opp ned på det vitenskapelige verdensbildet, og som er mest kjent som Darwins evolusjonsteori (1).
Situasjonen over illustrerer Darwins grunnleggende tanke og provokasjon i et nøtteskall, og logikken er ganske enkel: En arvelig egenskap sprer seg i befolkningen avhengig av effekten den har på forplantningsevnen. En egenskap som fører til tidlig død vil nødvendigvis ha dårlig evne til å spre seg, mens en egenskap som gir langt liv og mye avkom, raskt vil forplante seg i befolkningen. Om en egenskap er fordelaktig eller ikke, er derimot fullstendig avhengig av det omkringliggende miljøet. En dødelig egenskap i dag kan være morgendagens vinner, og selv små forandringer i omgivelsene kan gi store effekter på overlevelsesevnen.
Denne vekselvirkningen mellom arv og miljø representerer fundamentet for Darwins evolusjonsteori. Men når grunnprinsippet er så enkelt, hvorfor er darwinismen da så provoserende? Problemet oppstår når vi anvender darwinistisk logikk på problemstillinger hvor forståelsen er basert på tradisjonelle lineære tankerekker (2). Spesielt kjent er selvsagt konflikten mellom Darwins og Bibelens forklaring på artenes opprinnelse, men det logiske gapet sees også i mer aktuelle problemstillinger.
Vår kamp mot patogene mikroorganismer er i stor utstrekning basert på en lineær tankerekke: Bakterier forårsaker sykdom, derfor dreper vi dem med antibiotika – og pasienten blir frisk. Slik har vi også oppdratt pasientene til å tenke. Darwinistisk logikk forutser derimot noen helt andre og langt mindre positive konsekvenser av vår behandlingsiver. Industriell produksjon og anvendelse av antibiotika innebærer i dette perspektivet en radikal endring i det mikrobiologiske miljø, som i henhold til evolusjonsteorien selekterer for nye egenskaper. Resultatet er en av vår tids største helseproblemer og kjennes som resistensutvikling og utradering av den normale bakterieflora (3).
Men selv om gamle tankerekker og forskrivningsvaner kan være vanskelig å bryte, er det i dag få som blir provosert av at man anvender darwinistiske modeller for å forstå bakterier. De dyptliggende problemene starter først når vi trekker våre egenskaper og vår eksistens inn i dette perspektivet.
Et studie som sammenlikner levealder hos primater er et spennende eksempel (4): Hypotesen er at forskjellen i levealder mellom menn og kvinner henger sammen med hvem som tar seg av barna, og den underliggende logikken er at det å leve lenge er en egenskap med størst reproduktivt potensial hos det omsorgsfulle kjønn. Sagt på en annen måte – hvis det er hunnen som tar seg av barna, vil de ha mer utbytte av at mor lever lenge enn at far gjør det. Darwinistisk logikk taler dermed for at det omsorgsfulle kjønn lever lengst, og studien av primater med henholdsvis maternelle og paternelle omsorgsfigurer underbygger nettopp dette (4). Anvendt på oss mennesker antyder denne modellen at delt forelderansvar gjennom mange generasjoner vil føre til utjevning av kjønnsforskjellen i levealder. Darwinistisk tankegang passer i dette tilfellet godt med dagens likestillingspolitikk.
Så politisk korrekt var ikke min uforvarende anvendelse av darwinistisk logikk på hjertesyke barn. For hvordan skal vi forholde oss til at vår iherdige streben for å berge liv vil føre til mer sykdom i befolkningen? Hva skal vi da gjøre med de medfødt syke? Her møter vår lineære behandlingslogikk et dilemma av uante dimensjoner, og spøkelset jeg hadde manet frem hos foreleseren, bærer eugenikkens fryktinngytende skikkelse (5). Med bilde av steriliseringslover og konsentrasjonsleirer i bakhodet hadde han unektelig all grunn til å bli sint, og han er langt fra den første som har satt likhetstegn mellom Darwins tanker og såkalt rasehygiene (6).
Men her ligger også den logiske bristen. Darwins evolusjonsteori beskriver den basale sammenhengen mellom arv og miljø, den forklarer utviklingsmønstre og kan brukes til å forutsi konsekvenser av miljøendringer. Den sier for eksempel at arvelige hjertelidelser vil bli mer vanlig i et miljø som reparerer dem, at overforbruk av antibiotika vil fremskynde resistensutvikling, eller at diabetes vil øke i hyppighet når insulin er konsumvare. Men evolusjonsteorien sier ingenting om at individer med avvikende trekk bør elimineres. Tvert imot, dette prinsippet er uavhengig av hva vi til enhver tid måtte oppfatte som sykdom eller uønskede egenskaper.
Begrepet «survival of the fittest», som Darwin overtok fra sosiologen Herbert Spencer (2), innebærer i all enkelhet at egenskaper som fremmer reproduksjon, forplanter seg gjennom generasjonene. En egenskaps «fitness» eller tilpassethet defineres utelukkende ut fra dens bidrag til forplantningsprosessen, og er i prinsippet uavhengig av kulturelt betingede normer for normalt og unormalt. En egenskaps tilpassethet måles i antall etterkommere med samme egenskap, enten dette innebærer hjertefeil, kortvoksthet, vinger, finner eller kropp som en supermodell. Alt avhenger av det omkringliggende miljøet.
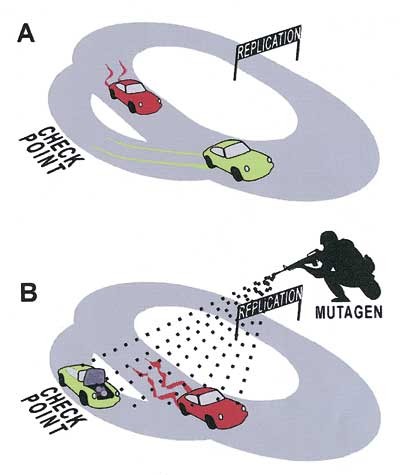
Figur 1 Cellesyklus Grand Prix og effekten av motsatte reparasjonsstrategier i ulike miljøer. Team 1 (grønn) stopper alltid for reparasjon når det oppstår en skade, mens Team 2 (rød) overser alle varsellamper. Team 1 vinner under normale forhold (A) fordi de alltid har et feilfritt kjøretøy, mens Team 2 hele tiden får nye skader. I det tøffere miljøet (B) akkumulerer begge kjøretøyene skader raskere enn de kan repareres, og Team 1 blir fanget i sjekkpunktet. Team 2 derimot humper av gårde i sitt skranglete kjøretøy med et rimelig håp om å nå mållinjen. Denne enkle analysen av reparasjonsstrategier gir dermed en forklaring på det tilsynelatende paradokset at mutagene miljøer favoriserer tap av DNA-reparasjonsgener. Gjengitt fra Breivik (16)
Det er en grov og utbredt misforståelse at evolusjonsteorien rettferdiggjør eliminering av svake individer, og denne uheldige koblingen understrekes ved at «survival of the fittest» på norsk oversettes med «den sterkestes rett». Darwinistisk tenkning settes på den måten opp mot humanismen, og fremstår som et verdisyn man velger å forholde seg til på ideologisk grunnlag. Slik er det ikke. Evolusjonsteorien representerer en meget velfundert og fundamental vitenskapelig modell med direkte relasjoner til grunnleggende prinsipper innen kjemi, fysikk og informasjonsteori (7). Den er som tyngdeloven deskriptiv og predikerende, ikke normativ – og Newtons oppdagelser er da vitterlig ikke noe argument for å dytte mennesker over ende, selv om de skulle være aldri så ustø.
Ikke stopp for reparasjoner i en krigssone
Evolusjonsteorien kan i dag forklares ut i fra DNA-molekylets struktur og dynamikk (8, 9). Dette perspektivet innebærer et nytt og spennende grunnlag for biomedisinsk forståelse, og ved Seksjon for Immunterapi ved Radiumhospitalet har vi de siste årene anvendt denne forståelsen for å belyse en av kreftforskningens mest grunnleggende problemstillinger.
Kreftceller er genetisk ustabile (10). For hver celledeling får de et stort antall mutasjoner, og slike genetiske endringer kan forklare kreftcellenes irregulære morfologi og vekstmønster. Entusiasmen var derfor stor da man i begynnelsen av 1990-årene fant at genetisk ustabilitet skyldtes mutasjoner i gener som regulerer reparasjon av andre gener (11). Ustabiliteten skyldtes logisk nok mangelfull DNA-reparasjon, og man kunne dermed forklare kreftutvikling med at feil i reparasjonsgener utløser en kaskade av skader i andre gener (12).
Men det er ett grunnleggende problem med denne lineære modellen: Kreftutvikling innebærer at kreftcellen formerer seg og fortrenger de normale cellene i kroppen. Men hvorfor skal en celle som ikke er i stand til å reparere skader, vokse raskere enn en tilsvarende celle som kan reparere sine skader? Den mest vanlige forklaringen er blitt at ustabile celler er mer tilpasningsdyktige, og at de mange mutasjonene hjelper cellene til å overleve (13). Dette er en tiltalende modell, med mange celebre tilhengere. Men med bakgrunn i den molekylære forståelsen av evolusjonsteorien har Gustav Gaudernack og jeg hevdet at den er basert på minst to logiske feilkoblinger.
For det første betyr fravær av reparasjon at cellenes DNA-molekyler utsettes for en haglstorm av mutasjoner. Disse mutasjonene rammer mer eller mindre tilfeldig. Risikoen for at en slik mutasjon ødelegger viktige gener, er mye større enn sjansen for en mutasjon med positiv effekt på celledelingen. Problemstillingen har en fundamental statistisk og termodynamisk basis, men kan enkelt illustreres med Fleksnes-metoden for å reparere defekte TV-apparater. Det vil si å slå og sparke i apparatet til bildet blir klart. En slik strategi kan gå bra én gang, kanskje to, men på lengre sikt vil summen av skader være destruktiv. I sum er det derfor vanskelig å påstå at genetisk ustabilitet er positivt for cellen, men fordi det bare er de positive mutasjonene som overlever, har mange latt seg forlede til å tro at ustabiliteten har en mening (14).
Et annet problem med dette perspektivet er litt vanskeligere å forklare, men henger sammen med grunnleggende forståelse av evolusjon. Modellen over sier at celler blir selektert fordi de muterer, men strengt tatt er det ikke celler som selekteres. Celler, organismer og arter er kun midlertidige konstruksjoner – det er den molekylære informasjonen i genene som kopieres og utvikler seg gjennom generasjonene (9). Ofte kan det være mest praktisk å snakke om seleksjon av celler eller organismer. Men når problemstillingen handler om gener som påvirker andre gener, blir det svært viktig å holde tungen rett i munnen.
Vi har derfor hevdet at genetisk ustabilitet krever en stringent molekylær forståelse av Darwins evolusjonsteori (15). Denne måten å se biologien på setter DNA-molekylet og den genetiske informasjonen i begivenhetenes sentrum, og modellen kalles gjerne «det egoistiske gen» eller «the selfish gene» etter Richard Dawkins’ populærvitenskapelige bok om temaet (9). Istedenfor å gruble over kreftcellenes besynderlige oppførsel, valgte vi derfor reparasjonsgenene som subjekt for problemstillingen, og spurte hvorfor disse genene kopierer seg så dårlig i kreftcellene. Problemet kokte på den måten ned til en vurdering av fordeler og ulemper ved å reparere gener, og for å illustrere logikken kan man sammenlikne reparasjonsgenene med strategier på en racerbane (fig 1) (16).
Hvor ofte man skal kontrollere og reparere et kjøretøy er en vanskelig avveiing, og bilfabrikantene bruker millioner på å analysere slike vurderinger. Sparsomt vedlikehold øker risikoen for havari, mens hyppige kontroller koster både tid og penger. Hvilken strategi man skal velge, avhenger av hvor man kjører, og den intuitive tanken er at tøffe forhold krever hyppig vedlikehold. Men slik er det nødvendigvis ikke. For hva om løypa er helt ekstrem? Hva om racet for eksempel går igjennom en krigssone hvor geriljasoldater beskyter kjøretøyet? Å stoppe for å reparere vil da være en dødelig strategi. Vår noe overraskende konklusjon ble derfor at selv om tøffe miljøer øker behovet for DNA-reparasjon, øker de også fordelen med å la det være.
Mutagene miljøer kunne dermed forklare hvorfor reparasjonsgenene blir borte, og i litteraturen fant vi støtte for at genetisk ustabilitet har sammenheng med kreftsvulstens omgivelser (17). Spesielt interessante var genetiske analyser som viser at høyresidige og venstresidige tykktarmssvulster er markant forskjellige. På høyre side forsvinner genene som reparerer små uregelmessigheter i repeterende DNA-sekvenser, mens det på venstre side er mekanismene som regulerer kromosomdeling og kromosomstruktur som går tapt.
Disse observasjonene holdt vi så opp mot epidemiologiske studier som viser at høyre- og venstresidig tykktarmskreft har sammenheng med ulike miljøfaktorer som gallesyrer, kosthold og røyking. Helt i tråd med våre antakelser var det sammenheng mellom kjemiske stoffer i miljøet og de reparasjonsmekanismene som var gått tapt i svulsten. Etter flere års systematisering av informasjon fremsatte vi derfor en hypotese om at miljøet styrer kreftutviklingen ved å favorisere tap av ulike reparasjonsgener (17).
Denne darwinistiske modellen ble raskt satt på prøve av en av kreftforskningens mest anerkjente forskergrupper, og treffsikkerheten var oppsiktsvekkende (18). Celler som ble utsatt for mutagene stoffer, døde hvis de hadde intakte reparasjonsmekanismer, mens de som vokste videre, manglet reparasjonsgener og var genetisk ustabile. Hvilken type ustabilitet som utviklet seg, var avhengig av miljøet cellen ble utsatt for. Arbeidet bekreftet dermed vår hypotese om darwinistisk samspill mellom gener og miljø i kreftutvikling.
Denne modellen har også betydning for hvordan man forstår ulike behandlingsformer, for kreftbehandling handler i stor utstrekning om å påføre kreftcellene dødelige skader. Tidligere trodde man at cellenes evne til reparasjon var en hindring for effektiv terapi, men vår darwinistiske modell snur dette på hodet (16). Skal man drepe kreftceller, bør man nettopp lure dem til å stoppe opp og åpne panseret (fig 1).
Darwins informasjonsteori
Denne problemstillingen fra kreftforskningen innebærer altså nok et eksempel hvor darwinistisk logikk snur forståelsen av levende systemer på hodet. Ting som formerer seg, har en dynamikk som innebærer kontinuerlige vekselvirkninger med det omkringliggende miljøet, og tilsynelatende logiske forklaringsmodeller basert på lineære tankerekker bryter sammen når det er selve livet som skal analyseres.
Denne darwinistiske forståelsen av fenomenet liv har også et utvidet perspektiv. Om man velger en biologisk eller en kulturell tilnærming til tilværelsen, handler livet alltid om overføring av informasjon – enten det er gener, nerveimpulser, bokstaver, lydbølger, bilder eller biter. Innen det vitenskapelige miljø er det i dag en gryende oppfatning av at evolusjonsteorien beskriver et dynamisk prinsipp som gjelder all informasjon, ikke bare den som er skrevet som DNA-molekyler. Det blir dermed like dekkende å snakke om en informasjonsteori som en evolusjonsteori, og i dette perspektivet har Darwins vitenskapelige revolusjon bare så vidt begynt (2, 19).
I denne utviklingen står vitenskapelig medisin i en særstilling, midt mellom biologien og kulturen. Innen genetikken ser man allerede at grensen mellom bioteknologi og informasjonsteknologi flyter over i hverandre. I nevropsykologien studerer man samspillet mellom kulturell og biologisk informasjon på stadig mer raffinerte måter. Samfunnsmedisinen møter en verden hvor vitenskapen om liv og informasjon i økende grad styrer utviklingen, og medisinens psykosomatiske dilemma nærmest skriker etter en vitenskapelig forståelse av samspillet mellom tanker og gener.
Medisinens tverrfaglighet representerer derfor et spennende utgangspunkt for moderne darwinistisk forståelse, men det medisinske miljø har i liten grad tatt innover seg Darwins revolusjonerende tanker. Mange oppfatter fremdeles darwinistisk logikk som en trussel mot medisinens kristen-humanistiske dogmer, og denne trusselen er i høyeste grad reell for dem som misliker nytenkning og provoserende spørsmål:
For hvorfor er det etisk høyverdig å bruke avansert medisinsk teknologi til å selektere sykdomsgener, mens seleksjon av gener med positive effekter er moralsk forkastelig? Og hvem har ansvaret når vår hellige behandlingsiver skaper mer sykdom?
Mitt arbeid for å formidle vitenskapelig forståelse for gener og liv finansieres av Norges forskningsråd.
