Sykdommer knyttet til fødeinntak og kroppsvekt forekommer hyppig og har store personlige og samfunnsmessige konsekvenser. Grunnleggende innsikt i mekanismene som kontrollerer sult og metthet er nødvendig for forståelsen av slike lidelser, ikke minst som grunnlag for behandling. En modell for den biologiske fødeinntaksreguleringen blir her beskrevet.
Kunnskapen om appetittens biologi har utviklet seg betydelig de siste 15 årene, og en del aspekter er tidligere omtalt i Tidsskriftet (1 – 3). Denne fremstillingen tar sikte på å gi et samlende overblikk over feltet.
En homøostatisk modell
Hvor mye og hvor ofte vi spiser, avgjøres av en rekke interne og eksterne faktorer, og årsakene til sykelige endringer i kroppsvekten må som oftest tilskrives sammensatte vekselvirkninger mellom biologiske, psykologiske og miljømessige forhold. Regnestykket som må gå opp for at en konstant kroppsvekt skal kunne opprettholdes, er likevel påfallende ukomplisert: Energiinntaket og energiforbruket må være av samme størrelse. Forestillingen om at kroppen aktivt regulerer fettlagerets stabilitet støttes av empiriske studier og har klare paralleller til andre kjente, homøostatiske kontrollsystemer (4). Ifølge denne tankegangen vil kroppsvekten til enhver tid forsvares mot avvik fra en gitt terskelverdi, interessant nok nærmest uavhengig av hva som forårsaket disse endringene. Mekanismene opererer likevel asymmetrisk, ved at energiunderskudd forsvares sterkere enn energioverskudd. Det var sikkert fordelaktig i våre forfedres kalorimagre miljø, men klart ugunstig med dagens spisekart (5). Regulering av kroppsvekten via kontroll av fødeinntaket kommer i stand ved et samspill mellom hjernen og de perifere vev.
Sentral regulering
Hypothalamus har vært tillagt funksjoner i kroppsvektreguleringen siden midten av 1800-tallet. Da begynte man å sammenholde kliniske observasjoner av pasienter med adipositas og kakeksi med patologisk-anatomiske undersøkelser av hjernen (6). Det var likevel først i 1940- og 50-årene, med arbeidene til Hetherington & Ranson (7) og Anand & Brobeck (8), at man fikk en tilfredsstillende teoretisk forståelse av den sentrale reguleringen av fødeinntaket. Den såkalte tosenterhypotesen forklarer sult og metthet ved tilstedeværelsen av et tonisk aktivt sultsenter i laterale hypothalamus, som hemmes av et måltidsaktivt metthetssenter i ventromediale hypothalamus (9). Denne klassiske teorien har riktignok overlevd lenge i lærebøkene, men representerer en utdatert overforenkling. I dag vektlegges i større grad betydningen av arcuatuskjernen og dens rolle som mottaksstasjon for sirkulerende hormoner. Det er identifisert to distinkte nevronpopulasjoner i denne kjernen, med et yin-yang-liknende forhold til hverandre: NPY/AGRP-cellene og POMC/CART-cellene. Disse igjen aktiverer nervekretser som befordrer henholdsvis anabole og katabole atferdsmønstre (fig 1). Det komplekse samspillet mellom nevropeptidene som inngår i disse kretsene, er under intens utforskning. Man har identifisert mutasjoner i signalveiene som grunnlag for humane, monogene overvektstilstander, der den såkalte MC4R-defekten forekommer klart hyppigst (1). MC4R (melanokortin-4-reseptor) stimuleres normalt av α-melanocyttstimulerende hormon (α-MSH) fra POMC/CART-cellene og videreformidler informasjon som gir metthetsfølelse. Faller dens funksjon bort (defekten nedarves autosomalt dominant), vil det gi seg til kjenne som hyperfagi (10).
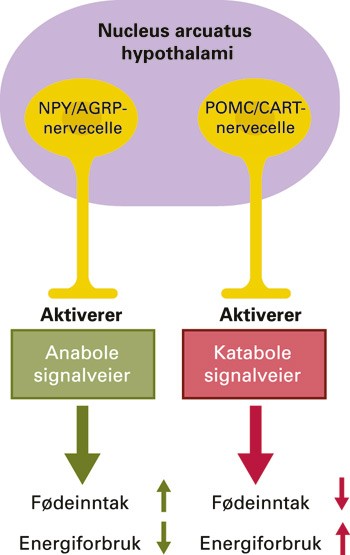
Figur 1 Arcuatushypotesen. Nevronpopulasjonene i arcuatuskjernen i hypothalamus er fremstilt skjematisk. Oreksigener (stoffer som fremmer fødeinntaket) stimulerer NPY/AGRP-signalveien og/eller hemmer POMC/CART-signalveien (eksempelvis ghrelin), mens anoreksigener (stoffer som hemmer fødeinntaket) stimulerer POMC/CART-signalveien og/eller hemmer NPY/AGRP-signalveien (eksempelvis leptin). NPY = nevropeptid Y, AGRP = agoutirelatert peptid, POMC = proopiomelanokortin, CART = kokain- og amfetaminregulert transkript
Hypothalamus har utstrakte, resiprokale forbindelser innad i sentralnervesystemet. Den kan dermed både påvirke og påvirkes av en lang rekke andre strukturer og fungere som en slags integreringssentral eller koordineringssete for matrelatert atferd. Særlig interessant er forholdet til hjernens belønningsbaner. Lystrelaterte impulser kan overstyre signalene knyttet til kroppsvektshomøostasen («dessertmage»-fenomenet), og slik sett er det visse likheter mellom spising og bruk av rusmidler (11).
Perifer regulering
Et utall biologiske signaler påvirker fødeinntaket, og flere periferfysiologiske forklaringsmodeller for sult og metthet har vært foreslått. Tanken om at sult skyldes underskudd av energi i bestemte kjemiske eller fysiske former og at metthet oppstår når dette underskuddet dekkes inn, har lenge stått sentralt, og ligger bak de såkalte deplesjon/replesjon-modellene: Lipostathypotesen, glukostathypotesen, aminostathypotesen og termostathypotesen. I dag synes en inndeling i langsiktige og kortsiktige mekanismer mer hensiktsmessig (12), og disse utgjøres for en stor del av henholdsvis fettvev-hjerne-aksen og tarm-hjerne-aksen.
Fettvev-hjerne-aksen
Kennedy postulerte i 1953 at størrelsen av kroppens fettdepot kan sanses og at denne informasjonen inngår i reguleringen av kroppsvekten (13). Informasjonsstrømmens koblingsrute mellom fettvev og hjerne danner en negativ tilbakemeldingssløyfe: En kvantitativ reduksjon av fettmassen fører til stimulering av et energikonserverende atferdsmønster, mens en økning fører til hemming av slik atferd. I 1973 viste Coleman at signalet i fettvev-hjerne-aksen er humoralt mediert (14). Hormonene som tar del i reguleringen kalles gjerne fedmehormoner, og generelt gjelder at deres nivå stiger proporsjonalt med fettmassen. Aktiviteten er «kronisk» ved at hormonnivået til enhver tid monitorerer og reflekterer den eksisterende mengde kroppsfett. Denne egenskapen gjør fettvevet i stand til å utøve en langsiktig regulering av fødeinntaket.
Det mest sentrale fedmehormonet vi kjenner til ble isolert av Friedmans gruppe i 1994 og navngitt leptin («leptos» betyr slank) (15). Leptinet og dets effekter blir i en omfattende mengde litteratur utpekt som viktigste enkeltstående bidrag til det kroppsvektregulerende systemet. Oppdagelsen av leptin førte til stor optimisme: Hadde man endelig funnet det magiske stoffet som overvektige manglet og som gjorde dem fete? Vi vet i dag at leptinmangel (genetisk betinget) er uhyre sjelden, men når det forekommer, kan tilstanden behandles kausalt – gjennom leptintilførsel – med strålende resultater. Ved vanlige fedmetilstander er derimot leptinnivået høyt (proporsjonalt med kroppsmasseindeks), og det kan synes å foreligge resistens mot sirkulerende leptin (16). Eventuell farmakoterapi må derfor rettes mot å bedre denne resistenstilstanden (17).
Fettvevet betraktes i dag som et aktivt endokrint organ, som skiller ut flere stoffer med liknende effekter som leptin (såkalte adipokiner), inkludert resistin og adiponektin (2). Fettvevets omfang, spesielt mengden innvollsfett, reflekteres også i insulinnivået. Foruten de velkjente, metabolske effektene i perifere vev, er det holdepunkter for at insulin regulerer kroppsvekten på samme måte som leptin – ved å redusere fødeinntaket og øke energiforbruket – via insulinreseptorer i hypothalamus (18).
Tarm-hjerne-aksen
Både sult og metthet projiseres ofte til magen, og at gastrointestinaltrakten bidrar til fødeinntaksreguleringen har vært kjent lenge. Hvor omfattende og komplisert dette bidraget er, har man begynt å ane konturene av først i den senere tid. Mage-tarm-kanalen innehar et velutviklet nervenett – det enteriske nervesystem (ENS) – med omtrent like mange nerveceller som ryggmargen (19). Denne «second brain» (20) signaliserer i større grad til hjernen enn omvendt, eksemplifisert ved at hele 80 – 90 % av fibrene i vagusnerven er afferente. Et utall enteroendokrine celler sikrer dessuten at hjernen også informeres om forholdene i tarmen via blodbanen, og utgjør derved en humoral tarm-hjerne-akse.
Allerede i 1912 foreslo Cannon og Washburn at sult skyldes spesielt sterke sammentrekninger av den tomme magesekkens vegg – sultkontraksjoner (21). Cannon hadde merket seg at det rumlet i magen når han var sulten, og satte ned en ballong i ventrikkelen til sin assistent Washburn for å belyse denne sammenhengen nærmere. I mange år var teorien om sultkontraksjonene nærmest enerådende som periferfysiologisk forklaringsmodell for fødeinntaksreguleringen. Med mer moderne metoder har man også funnet en sammenheng mellom sultfølelse og fastemotorikk i tarmkanalen (22), men som hovedforklaring på sult er teorien i dag utdatert. Mennesker som har fått fjernet sin ventrikkel, blir også sultne.
Fylling av magesekken som årsak til metthet er velkjent. Ventrikkelens evne til å signalisere metthet har vært knyttet til fundusdelens akkommodasjonsmekanisme. Når veggen relakseres under oppfylling, vil slimhinnefoldene (rugae gastricae) brette seg ut og blottlegge tidligere skjulte reseptorelementer, som derved blir aktivert (23). Mennesker med svekket ventrikkelakkommodasjon, eksempelvis pasienter med funksjonell dyspepsi, kjennetegnes imidlertid ved nettopp tidlig metthet, stinnhet og oppfylthet ved inntak av mat, ofte ledsaget av en utvidelse av magesekkens antrum (24). Kanskje er det slik at fylling av fundusdelen gir en behagelig metthetsfølelse, mens fylling av antrumdelen gir en ubehagelig metthetsfølelse?
Gastrointestinale metthetshormoner har vært i søkelyset helt siden Smith & Gibbs i 1973 tilla kolecystokinin (CCK) hemmende effekt på fødeinntaket hos rotter (25). Kolecystokinin anses i dag som arketypen for en lang liste av foreslåtte metthetspeptider, inkludert polypeptid YY (PYY), glukagonliknende peptid-1 og -2 (GLP-1 og GLP-2), oxyntomodulin (OXM), gastrisk inhibitorisk peptid (GIP), somatostatin, enterostatin, nevrotensin og bombesinliknende peptider fra tarmen samt pankreatisk polypeptid (PP), amylin og glukagon fra bukspyttkjertelen (26). Metthetspeptidene deltar først og fremst i den kortsiktige reguleringen av fødeinntaket, men enkelte av dem har nok også langsiktige virkninger. Hormonenes rolle i medikamentell behandling av overvekt er under utforskning (27).
Mage-tarm-kanalens hovedoppgave – nedbrytning av mat og absorpsjon av næring – avhenger av at viktige støttefunksjoner, som sekresjon, motilitet og sirkulasjon, koordineres nøyaktig i tid og rom. Det kan synes som om den gastrointestinale reguleringen av fødeinntaket primært er utviklet for å ivareta slike hensyn. Således er det logisk at de fleste signalene fra tarmen har karakter av negative tilbakemeldingssløyfer (gir metthetsfølelse). Ghrelin representerer et interessant unntak («ghre» betyr vekst og «relin» betyr frisettelse): Oppdagelsen av at dette veksthormonstimulerende peptidet fra magesekken gir sultfølelse, har vakt berettiget oppsikt (28). Ghrelin og leptin virker mot hverandre på flere nivåer i signalveiene, og samspillet mellom dem er blitt sammenliknet med dansen tango (29). Ghrelinnivået er lavt ved vanlige fedmetilstander og inverst korrelert med kroppsmasseindeks. Dette mønsteret ses riktignok ikke ved Prader-Willis syndrom – en genetisk betinget tilstand kjennetegnet ved svær appetitt og fedme – der meget høye nivåer av ghrelin er rapportert (30). Blokkering eller stimulering av ghrelinsignalet er interessant for fremtidig farmakoterapi, også for andre sykdommer enn lidelser i kroppsvektreguleringen, som hjertesvikt og osteoporose (31).
Bariatrisk kirurgi (operativ behandling av sykelig overvekt) er nylig omtalt i et temanummer av Tidsskriftet (nr. 1/2007). Inngrepene er basert på innførsel av mekanisk restriksjon og malabsorpsjon, men har etter hvert også vist seg å indusere vesentlige endringer i den gastrointestinale hormonsekresjonen. Eksempelvis fører biliopankreatisk avledning med duodenal omkobling til fall i ghrelinkonsentrasjonen (fordi store deler av ventrikkelen, der ghrelin syntetiseres, skjæres vekk) og økning av nivåene av polypeptid YY og glukagonliknende peptid-1 (fordi malabsorbert føde kommer i kontakt med endokrine celler i distale ileum). Dette kan tenkes å bidra til den gunstige vektreduserende og antidiabetiske effekten ved slike inngrep (32). Det er likevel ikke avklart hvor stabile disse humorale endringene er (33).
Samspill
Mens fedmehormonene utøver sin effekt i hypothalamus og gir en langsiktig kontroll av kroppsvekten, konvergerer signalene fra mage-tarm-kanalen, både nevrale (vagusnerven) og humorale (metthetspeptider), hovedsaklig i hjernestammen og bidrar her til en kortsiktig regulering av det enkelte måltid (34). Unntakene er sulthormonet ghrelin og en del av metthetspeptidene, som virker direkte i arcuatuskjernen. Hjernestammen synes å ha tilstrekkelig sensorisk, integrerende og motorisk kapasitet til å kunne utøve måltidsregulering nærmest uavhengig av overordnede hjernesentre. Dette ble blant annet påvist av Grill & medarbeidere i studier av decerebrerte rotter (35). En «justering» fra hypothalamus er likevel nødvendig for at kroppsvekten skal holde seg stabil på sikt. Forenklet kan vi si at det langsiktige systemet eller kroppsvektsreguleringen overstyrer og modulerer responsterskelen til det kortsiktige systemet eller måltidsreguleringen (36).
Samvirket mellom hjernen, fettvevet og mage-tarm-kanalen er utsatt for en rekke påvirkninger utenfra og kan dermed betraktes som en slags homøostatisk grunnstruktur i en større forklaringsmodell for sult og metthet, der ikke bare andre biologiske, men også psykologiske og miljømessige forklaringer inngår.
Konklusjon
Sult og metthet blir langt på vei bestemt av fysiologiske mekanismer som ivaretar stabiliteten i kroppens indre miljø. I denne artikkelen er hovedtrekkene i den biologiske fødeinntaksreguleringen belyst ved å innføre et skille mellom sentrale og perifere systemer, og mellom kortsiktige og langsiktige mekanismer. Slike inndelinger er selvsagt kunstige og innebærer en fare for overforenkling, men kan like fullt være nyttige innfallsvinkler til et stadig mer uoversiktlig tema i medisinen.
