Karakteristisk for kols er en luftveisobstruksjon som ikke er fullstendig reversibel. De siste 40 år er spirometri – med lungefunksjonsmålene forsert ekspiratorisk volum på ett sekund (FEV₁) og forsert vitalkapasitet (FVC) – blitt brukt til diagnostikk og klassifisering (1). Kols er en svært vanlig sykdom i Norge. Anslagsvis 200 000 mennesker er rammet, og det er ca. 30 000 nye tilfeller per år (2–4).
Røyking og andre miljøfaktorer er de dominerende risikofaktorene for utvikling av kols, og det er holdepunkter for at det er et dose-respons-forhold mellom mengde tobakksrøyk og grad av lungesykdom. Samtidig er det mye som tyder på at også genetiske faktorer er av betydning. For det første er det vist at kun 15 % av variabiliteten i FEV₁ kan tilskrives røyking (5) og at det er et mindretall av røykerne som utvikler sykdommen. For det andre er det påvist familiær opphopning av redusert lungefunksjon.
Denne artikkelen gir en oversikt over metoder for påvisning av genetisk disposisjon og en sammenfatning av nåværende kunnskap innenfor feltet genetikk og kols.
Materiale og metode
Grunnlaget for artikkelen er et ikke-systematisk litteratursøk i PubMed med et skjønnsmessig utvalg av artikler basert på forfatternes kunnskaper innen feltet og i tillegg erfaringer fra egne studier.
Genetiske begreper
Genetiske studier gjøres på DNA-nivå, mens ekspresjonsstudier (som gir informasjon om geners aktivitet) gjøres på RNA-nivå. Proteomikk er studien av proteiners struktur og funksjon. Funksjonell genomforskning (genomikk) beskriver geners og proteiners funksjoner og interaksjoner (fig 1). Det er identifisert mange millioner steder i genomet der enkeltnukleotider i DNA-sekvensen kan variere mellom to mennesker. Disse variantene kalles polymorfismer. Som markører i genetiske studier benyttes ofte enkeltbasevarianter (single nucleotide polymorphisms). De fleste har ingen funksjonell effekt, men noen kan påvirke disposisjon for sykdom.
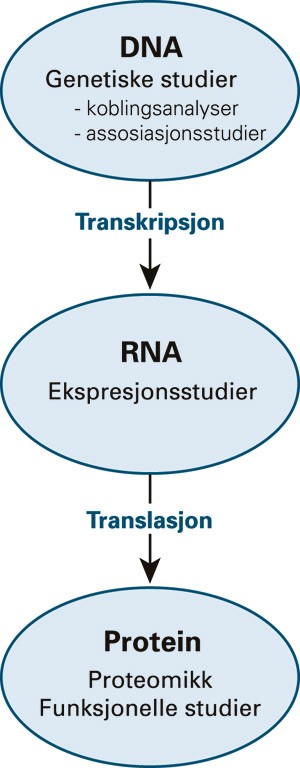
Figur 1 Genomforskning på ulike nivåer
Med genotype menes et individs genetiske sammensetning, mens fenotype beskriver fysiske karakteristika eller egenskaper hos et individ. Ulike fenotyper ved kols kan bl.a. bygge på symptomer, klinisk diagnose, lungefunksjonsmålinger, funn ved bildediagnostikk eller inflammasjonsmarkører (6).
Sykdommer påvirket av gener kan være monogene (mutasjon i ett enkeltgen), polygene (flere gener) eller multifaktorielle/komplekse (flere gener og miljøfaktorer). Kols er et typisk eksempel på en kompleks sykdom, med sannsynlig påvirkning fra en rekke gener og miljøfaktorer og interaksjoner mellom ulike gener og mellom gener og miljø (7).
Påvisning av genetisk disposisjon.
Familiestudier av arvelighet
Opphopning av en kompleks sykdom i familier er ofte første holdepunkt for at en sykdom kan ha en genetisk komponent. I familiestudier sammenliknes risikoen for sykdom hos slektninger til den sykdomsaffiserte med risikoen i den generelle populasjonen. Påvisning av økt risiko for sykdom hos familiemedlemmer kan skyldes at sykdommen er genetisk betinget, men det kan også skyldes opphopning av miljøfaktorer (f.eks. røyking) i familier. Ved tvillingstudier kan grad av arvelighet estimeres. Større overensstemmelse mellom eneggede tvillinger enn mellom toeggede tyder på arvelighet.
Familiebaserte koblingsstudier
Koblingsstudier benyttes for å finne regioner i genomet hvor genetiske determinanter for en sykdom er lokalisert. I disse studiene kan man anvende små familieenheter, f.eks. pasienter med foreldre, eller et større antall familiemedlemmer i familier der sykdommen opptrer (et slektstre). Dersom et gensegment opptrer hyppigere enn man skulle forvente hos de affiserte familiemedlemmene, tyder dette på at området kan være av betydning for sykdommen.
Assosiasjonsstudier med kandidatgener
Assosiasjonsstudier kan være både familie- og populasjonsbaserte, men gjøres oftest i pasient-kontroll-studier. Ved denne metoden er målet å lokalisere mulige enkeltbasevarianter i kandidatgener som kan påvirke utviklingen av sykdom. Kandidatgener kan ha kjent eller ukjent funksjon og kan velges ut fra genekspresjonsstudier, ut fra hypoteser omkring sykdomsmekanisme eller fra områder påvist ved koblingsstudier. Enkeltbasevariantene har flere varianter (alleler), og man sammenlikner frekvensen av variantene i pasient- og kontrollgruppen. Signifikant forskjell i allelfrekvens mellom pasienter og kontrollpersoner kan ha flere årsaker (8): en enkeltbasevariant disponerer for sykdom, enkeltbasevarianten er lokalisert i nærheten av og arves sammen med sykdomsgenet (koblingsulikevekt) eller det kan være falskt positive assosiasjoner grunnet skjevhet (bias) (f.eks. etniske forskjeller), for lite utvalg eller statistisk tilfeldighet.
I tillegg til analyse av individuelle enkeltbasevarianter kan også koblede nærliggende enkeltbasevarianter på et kromosom (haplotyper) undersøkes med tanke på assosiasjon.
Genomvide assosiasjonsstudier
Nye teknologiske fremskritt har muliggjort genomvide assosiasjonsstudier (GWAS) (9), med bruk av mange hundre tusen enkeltbasevarianter spredt utover genomet. Bakgrunnen for utviklingen er bl.a. det internasjonale HapMap-prosjektet (10), en database over vanlige enkeltbasevarianter i genomet, samt tilgjengelig teknologi som har gjort genotyping av store antall enkeltbasevarianter billigere. Med denne metoden undersøkes store deler av genomet, med mulighet for å påvise hittil ukjente kandidatgener for en rekke sykdommer (11). Begrensningene ved genomvide studier er bl.a. at antall analyser øker risikoen for falskt positive assosiasjoner og at polymorfismer må være forholdsvis hyppig forekommende i befolkningen for å kunne oppdages. Metoden er velegnet til å lete etter nye kandidatgener og kan dermed virke hypotesegenererende.
Genetisk disposisjon
Lungefunksjon
Lungefunksjon målt ved spirometri er den mest brukte fenotypen i genetiske studier av kols. En rekke befolkningsstudier i Norge (12, 13) har vist at 30–70 % av variasjonen i spirometriske variabler lar seg forklare ut fra kjønn, alder og kroppshøyde. Familiestudier fra Frankrike (14) og tvillingstudier fra Sverige (15) har vist at arv representerer nærmere 20–30 % av variasjonen i lungefunksjon etter justering for alder, kjønn, høyde og røykevaner. Koblingsanalyser med lungefunksjonsmål er gjort i flere familiestudier i USA. Noen grad av kobling for FEV₁ og FEV₁/FVC er funnet til kromosom 6, mens FVC var koblet til kromosom 21 (16). I en annen studie fant man kobling mellom FEV₁/FVC og kromosom 4, mens FEV₁ og FVC var koblet til kromosom 18 (17). Så langt har det vært manglende reproduserbarhet av disse funnene.
Alfa-1-antitrypsinmangel
Alvorlig mangel på alfa-1-antitrypsin er en kjent genetisk risikofaktor for kols. Alfa-1-antitrypsin er et protein som kodes fra SERPINA1-genet på kromosom 14. De opprinnelig påviste variantene (18) ble benevnt etter deres elektroforetiske migrasjon: F (fast), M (medium), S (slow) og Z (very slow). Betydelig økt risiko for kols finnes hos røykere som er homozygote for proteaseinhibitorfenotype ZZ (forekomst 0,06 % i Norge). Samtidig er det noen ZZ-personer som ikke utvikler lungesykdom, noe som tyder på at andre gener og miljøfaktorer modifiserer effekten. Risiko ved fenotype MZ er grundig undersøkt. En befolkningsstudie fra Oslo viste at de som var heterozygote for Z ikke hadde mer obstruktiv sykdom enn de med fenotype MM (19), men dette er fortsatt uavklart (20).
Kols uten alfa-1-antitrypsinmangel
Kun en liten del av kolstilfellene skyldes alfa-1-antitrypsinmangel (19). Flere andre gener er trolig av betydning. En eldre tvillingstudie har vist betydelig familiær opphopning av hoste i noen familier av røykere (21). En annen studie viste at en familieanamnese med kronisk bronkitt og emfysem ga en oddsratio på 5,2 for at vedkommende selv hadde de samme sykdommene (22). Hos pasienter i Boston, USA, med tidlig debut av alvorlig kols hadde nære slektninger betydelig økt risiko for kronisk bronkitt og lav FEV₁-måling (23).
Det er foreløpig bare publisert én koblingsstudie for kvalitative og kvantitative fenotyper ved kols (24, 25). Mulige koblinger ble funnet til kromosom 22 for kronisk bronkitt hos røykere og til kromosom 12 for lav FEV₁-måling hos røykere. FEV₁ viste kobling til flere områder, mest uttalt til kromosom 8 og kromosom 1. FEV₁/FVC-ratio viste sterk grad av kobling til kromosom 2 og, i mindre grad, til kromosom 1. På den lange armen til kromosom 2 identifiserte man kandidatgenet SERPINE2 ved genekspresjonsstudier av lungevev.
Både familiebaserte studier og pasient-kontroll-studier har vist assosiasjon mellom SERPINE2 og kols (26), men en annen forskningsgruppe var ikke i stand til å bekrefte dette (27). Assosiasjonen ble deretter bekreftet i ytterligere to populasjoner – en familiestudie fra Europa/USA og en pasient-kontroll-studie fra Bergen (28). For fem av de 25 enkeltbasevariantene som ble testet var det en assosiasjon mellom SERPINE2-genet og kols i begge studiene. At funnet ble reprodusert i to store, uavhengige populasjoner, gir sterke holdepunkter for at SERPINE2 kan være et sykdomsgen. Hvilken rolle det eventuelt har i sykdomsutviklingen, er ennå ukjent.
I Bergen har vi utført assosiasjonsstudier med andre kandidatgener i det samme pasient-kontroll-materialet. I disse studiene testet vi enkeltbasevarianter i kandidatgenene ADAM33, LTA, ADRB2, EPHX1 og TNF. Ved analyse av en firedel av materialet fant vi en svak beskyttende effekt av en polymorfisme i EPHX1-genet (29), men ved analyse av hele materialet fant vi ingen overbevisende assosiasjon til noen av enkeltbasevariantene (30). Ved å skille pasienter og kontrollpersoner i mer ekstreme lungefunksjongrupper fant vi imidlertid assosiasjon til en enkeltbasevariant i TNF-genet (31).
Nylig ble den første genomvide assosiasjonsstudien for kols publisert. En av de undersøkte populasjonene var den nevnte pasient-kontroll-studien fra Bergen (32). Det ble identifisert to nye områder i genomet med assosiasjon til kols. CHRNA 3/5-genet ligger på kromosom 15 og koder for en nikotinreseptor. Dette genet har også vært assosiert med lungekreft i flere studier (33–35). Det er imidlertid uklart om dette virkelig er et sykdomsgen eller om det kun er involvert i nikotinavhengighet. HHIP-genet koder for «hedgehog»-interagerende protein og er lokalisert på kromosom 4. Man tror at dette genet kan være involvert i lungeutvikling, men funksjonen i kolssammenheng er ukjent. En annen genomvid studie har vist at HHIP er assosiert med lungefunksjon (36).
Manglende reproduserbarhet
I de fleste assosiasjonsstudier har man satt søkelys på mutasjoner i gener som påvirker proteaser/antiproteaser, antioksidanter/oksidanter, betennelsesreaksjoner, immunrespons og astma (ramme 1). Gjennomgående har den første publiserte studien vist en positiv assosiasjon, mens man i senere studier ikke har klart å reprodusere funnene (37). Det er flere årsaker til inkonsistente resultater og manglende reproduserbarhet (38). Et viktig problem har vært forskjellige pasientdefinisjoner (fenotypisk heterogenitet). I tillegg har definisjonen av kols variert over tid. Flere genetiske årsaker til samme sykdom (genetisk heterogenitet) er også en mulig forklaring. Dersom pasienter og kontrollpersoner kommer fra ulike populasjoner og har ulik genetisk bakgrunn, kan falskt positive sammenhenger oppstå – et fenomen kjent som populasjonsstratifisering. Lavt deltakerantall i studiene, statistiske utfordringer eller ufullstendig penetrans kan være andre årsaker.
Ramme 1
Kandidatgener som er blitt assosiert med kols, modifisert etter Silverman og medarbeidere (7)
Fremtidige forskningsretninger
Forskning på genetisk komplekse sykdommer er tverrfaglig og omfatter analyser av fenotyper og genotyper i aktuelle populasjoner med relevante statistiske metoder (fig 2). Denne forskningen kan bidra til oppdagelsen av nye markører for sykdommen, noe som kan være et verdifullt redskap både diagnostisk og prognostisk. Kunnskap om genetikk kan også gi innsikt i hittil ukjente sykdomsmekanismer og kan på sikt åpne for nye behandlingsmuligheter.
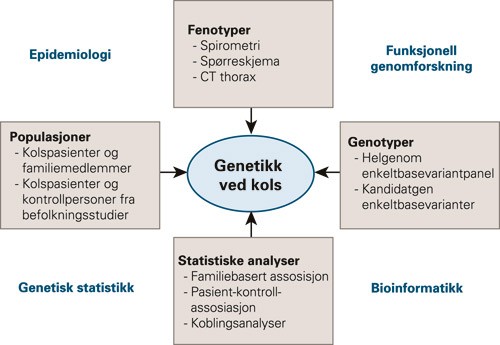
Figur 2 Tverrfaglig tilnærming til genetikk ved kols, modifisert etter Silverman og medarbeidere (7)
Teknologiske fremskritt gir stadig nye muligheter innenfor genetisk forskning. Fremover vil flere genomvide studier bli utført, og det å reprodusere positive funn i flere uavhengige befolkninger vil stå sentralt. Stor interesse knytter det seg også til fagfeltet proteomikk, der man håper å kunne måle proteiner i celler, vev og kroppsvæsker på samme måte som man måler RNA ved ekspresjonsstudier.
I klinisk forskning bør bedre standardisering av fenotypene og karakterisering av pasientene ha høy prioritet, slik at ulike studier blir sammenliknbare. Kols er en heterogen sykdom, og dersom et gen er viktig for en spesiell fenotype, f.eks. emfysem, finner man ikke nødvendigvis en assosiasjon dersom alle med en kolsdiagnose er blitt inkludert. Fremover bør man legge vekt på å sammenlikne genotyper med mer spesifikke fenotyper, f.eks. basert på CT-funn, andre lungefunksjonsmål, biomarkører eller fall i lungefunksjon over tid. Den genetiske bakgrunnen for nikotinavhengighet må undersøkes nærmere, det samme må interaksjoner mellom ulike gener og mellom gener og miljøfaktorer. Store internasjonale multisenterstudier med pasienter, familiemedlemmer og kontrollpersoner som følges over tid bør være idealet.
Genetiske studier av kols er i sin begynnelse, og mye er fortsatt ukjent. Forhåpentligvis vil videre forskning gi svar på hvilke gener som påvirker utviklingen av kols og identifisere de ulike genetiske varianter som er av betydning for sykdomsutviklingen.
